|
|
|
|
|
QB 北京大学李志远团队提出构建微生物铁互作网络的观点文章 |
|
|
论文标题:Forging the iron-net: Towards a quantitative understanding of microbial communities via siderophore-mediated interactions
期刊:Quantitative Biology
作者:Shaohua Gu, Jiqi Shao, Ruolin He, Guanyue Xiong, Zeyang Qu, Yuanzhe Shao, Linlong Yu, Di Zhang, Fanhao Wang, Ruichen Xu, Peng Guo, Ningbo Xi, Yinxiang Li, Yanzhao Wu, Zhong Wei, Zhiyuan Li
发表时间:10 January 2025
DOI:https://doi.org/10.1002/qub2.84
微信链接:点击此处阅读微信文章
铁元素是几乎所有生命形式必需的营养物质,对微生物的生长和代谢至关重要。然而,由于铁在自然环境中多以难溶的氧化铁形式存在,其生物可利用性极其有限。为了获取这种稀缺资源,微生物进化出了多种铁吸收机制,其中最主要的是通过分泌和吸收铁载体(siderophore)。铁载体是一类能够特异性螯合铁离子的小分子,被分泌到环境中与铁结合,而后被微生物膜表面的受体特异性识别并转运入细胞内。
不同微生物产生的铁载体结构各异,形成了复杂的化学多样性;而不同结构的铁载体具有对受体的高度特异性;于此同时,很多微生物通过在基因组中保持多条不同铁载体受体基因的方式,“窃取”其它微生物所制造的铁载体。综合起来,铁载体以其普遍性、多样性、特异性,介导了跨物种的竞争和博弈,在微生物群落中联系起一个复杂的铁互作网络,对理解和调控微生物群落具有重要意义。
近期,北京大学定量生物学中心李志远团队与南京农业大学韦中团队等在Quantitative Biology发表了题为"Forging the Iron-Net: Towards a Quantitative Understanding of Microbial Communities via Siderophore-Mediated Interactions"的观点文章。该文系统性地阐述了微生物铁载体网络的研究现状、理论框架和应用前景,提出通过构建“铁网”(Iron-Net)来理解和调控微生物群落的新思路。文章从分子机制、进化动力学到生态网络等多个层面,详细探讨了铁载体介导的微生物互作关系及其生态学意义。

全文概要
该文章从铁载体介导的铁互作网络探索了微生物群落;利用分子共进化预测铁载体介导的互作;铁互作网络中的微生物生态关系;次级代谢通过化学创新的进化意义这四个方面进行了探讨,并对微生物"铁网"构建的实践意义进行了讨论。
1. 通过铁载体介导的铁互作网络探索微生物群落
文章首先详细阐述了铁元素在微生物生存中的重要性和特殊性。作为地壳中含量最丰富的元素之一,铁元素却因其在含氧环境下易形成难溶性的氧化物而难以被微生物利用。研究发现,自然环境中的可利用铁浓度普遍远低于微生物生长所需:海洋中仅为10e-11-10e-10 mol/L,土壤中约为10e-18 mol/L,人体细胞外环境更是低至10e-24 mol/L,而微生物正常生长通常需要10e-7-10e-5 mol/L的铁浓度。即便在低pH或缺氧环境下,铁的可利用性仍然是制约微生物生长的关键因素(图1A)。
为了获取这种稀缺的铁资源,微生物进化出了多种获取机制,包括铁载体介导吸收、直接从宿主蛋白获取以及还原性铁吸收等途径。其中,铁载体介导的吸收机制最为普遍。铁载体是微生物分泌的一类具有强铁螯合能力的小分子,能与环境中的铁离子结合形成复合物,随后被微生物细胞特异性识别和吸收(图1A)。
铁载体展现出惊人的结构多样性。目前已实验确定的铁载体结构超过700种,而在自然产物库中可能存在数千种具有铁结合能力的结构。通过生物信息学分析发现,即使在研究较为充分的假单胞菌属中,仍有大量未被发现的铁载体结构存在。研究团队的初步分析表明,约60%的微生物基因组中至少包含一条铁载体合成途径。
铁载体与其识别受体之间存在高度特异的识别关系。革兰氏阴性和阳性细菌分别通过不同的机制识别和转运铁载体-铁复合物。前人研究发现,不同铁载体往往只能被特定的受体识别和吸收。例如,在铜绿假单胞菌中,三种不同菌株产生的不同pyoverdine(铁载体中的一大类)只能被各自的受体识别,几乎不存在交叉识别现象(图1B)。
然而,铁载体的“盗窃”现象在微生物界普遍存在。许多微生物能够利用其他物种产生的铁载体,这种现象不仅存在于亲缘关系接近的物种之间,还跨越了属、科甚至界的界限。例如,大肠杆菌不仅能利用自身产生的enterobactin,还能吸收其他多种微生物产生的铁载体。这种广泛的交叉利用现象得益于微生物基因组中通常携带多个识别受体(图1C)。
基于以上认识,作者提出了构建一个系统性的"铁网"的设想——如果能知道每个微生物制造和吸收什么铁载体,我们就能将各类微生物用铁相互作用联系起来,构成一张巨大的网络。铁元素的普遍必需性以及铁载体系统的广泛存在,确保了这一方法的普适性。同时,铁载体的多样性及其与受体间的特异性关系,为微生物群落的精确调控提供了可能。
图1. 铁载体介导的互作网络为微生物群落提供了机制性的了解和定向调控的可能。A. 铁载体-微生物关系概览。天然环境中的三价铁浓度远低于微生物生长需求(上左),而微生物产生铁载体来螯合和吸收铁(左下)。已知铁载体的化学多样性和一些结构的列子被展示在TMAP(Tree-MAP)视图中(右上)。图中还展示了革兰氏阴性和阳性菌中不同的铁载体吸收机制,突出了TonB依赖型受体(TonBDRs)和周质结合蛋白(PBPs)的分布情况。B. 不同铜绿假单胞菌中铁载体与特定受体的特异性识别关系。所示的三种铁载体可在铁载体信息数据库SIDERITE中查询。受体用凹槽旗形表示。C. 不同物种间铁载体的交叉利用。双箭头表示该物种既能产生又能利用该铁载体,对应的识别受体带有边缘;单箭头表示该物种只能利用(但不能产生)该铁载体,对应的识别受体无边缘。图中所有铁载体名称均可在铁载体信息数据库中查询。D. 微生物间铁载体介导的互作网络示意图。不同类型的铁载体及其在微生物膜上对应的识别受体(类型用不同颜色表示,且匹配的铁载体和受体是同一颜色)。微生物用圆角矩形表示,铁载体用彩色六边形表示,识别受体用带凹槽的旗形表示。识别受体可以吸收相同颜色的铁载体。黑色箭头表示产生关系,彩色箭头表示利用关系。后续图中使用相同的符号系统来说明铁载体互作。E. 利用不同铁载体调控微生物群落状态的示意图。
2. 从序列到生态——利用分子共进化预测铁载体介导的互作
在构建铁网的过程中,首要任务是对每个微生物进行“铁载体分型”,即确定它们能产生和利用哪些类型的铁载体。然而,这项工作面临着实验能力有限和生信分析复杂的双重挑战。特别是当一个基因组中存在多个识别受体时,我们往往难以区分哪些是用于识别自身产生的铁载体,哪些是用于“盗窃”其他微生物产生的铁载体。
作者最近发表于Science Advances的工作表明,基因共进化是应对这一难题的解题思路之一。这一思路的基础在于:铁载体的合成对微生物来说代价高昂。微生物主要通过两种途径合成铁载体:非核糖体肽合成酶(NRPS)途径和非NRPS铁载体(NIS)合成途径。其中NRPS途径负责约70%已知铁载体的合成,它需要庞大的酶系统,对细胞来说是巨大的投资。以假单胞菌为例,产生铁载体的菌株比非产生菌株的生长速率低3-10%(图2A)。
作者认为,在如此高昂的合成代价下,自然选择会推动铁载体与其对应识别受体之间的结构共进化——只有当铁载体和识别受体相匹配时,微生物才能有效地吸收铁离子,维持适应度。任何导致不匹配的突变都会使微生物承担合成代价而得不到收益,从而被选择所淘汰(图2B)。
基于这一认识,作者提出三种计算方法来预测铁载体-受体的配对关系:一是基于基因组邻近性,许多微生物的铁载体合成基因与其对应的识别受体基因在基因组上相距较近;二是利用基因序列中的协变信息,通过最大化共进化信号强度来识别匹配的铁载体-受体对;三是结合现在迅速发展的蛋白质结构预测和分子对接技术,在分子水平上预测互作关系(图2C)。
这些方法为从基因组序列预测微生物的铁互作行为提供了可能。作者设想了一个预测流程:输入基因组序列,通过预训练模型(该模型已从大量微生物序列数据中学习铁载体-受体的配对关系)进行分析,最终输出该微生物的“铁功能群”——它能生产和利用哪些类型的铁载体。这种“从序列到铁功能群”的智能分型方法,将帮助我们利用迅速增加的基因组数据来研究微生物世界的铁互作网络(图2D)。
图2. 基于基因组序列的数字化铁载体分型。A. 两种主要的铁载体合成途径概述:非核糖体肽合成酶(NRPS)途径(上)和非NRPS铁载体(NIS)合成途径(下)。约70%的微生物铁载体由NRPS途径合成,约30%由NIS途径合成。B. 铁载体和其识别受体之间结构共进化的选择压力示意图。C. 检测铁载体合成酶与识别受体共进化的三种计算方法:基因组邻近性分析;序列协变分析;结合分子对接的结构预测方法。D. 从微生物基因组预测铁功能群的可能流程。“铁功能群”可以用二元向量表示,用于识别每个菌株产生和利用的铁载体类型。
3. 铁互作网络——微生物生态博弈的游戏场
微生物在获取铁元素的过程中,形成了复杂有趣的生态互作关系。作者在假单胞属的接近2000菌株中观察到三类主要的铁获取策略:纯生产者(仅产生和利用自己的铁载体)、部分生产者(既产生自己的铁载体,也能利用他人的铁载体)和纯盗窃者(不产生但可利用他人的铁载体)(图3A)。有趣的是,不同生境和生活方式会偏好不同的铁获取策略,特别是在病原型生活方式中,群落中往往缺乏“部分生产者”这种策略。
为了系统分析两个微生物之间铁载体介导的复杂互作,作者提出了铁载体互作矩阵(SIM)的概念。这是一个2×2的矩阵,描述了两个菌株在“自受体”(识别和摄取自己生产的铁载体种类)和“盗受体”(识别和摄取自己不生产的铁载体种类)上的重叠关系(图3B)。矩阵的每个元素代表不同类型的互作:例如,若两个微生物的自受体类型相同,表示它们趋同生产相同的铁载体;反之则表示它们通过生产不同铁载体进行直接的铁竞争(图3C)。
铁载体的这种互作框架能够体现多个经典的生态学现象。例如,当两个微生物产生不同的铁载体且不能交叉利用时,就会出现“竞争排除”现象,即最终只有一方能够存活(图3D,1)。当一个纯盗窃者利用生产者的铁载体时,可能导致“公地悲剧”,即合作行为的崩溃(图3D,2)。在前人实验中,不产生铁载体的盗窃者株系确实能在多种条件下胜过生产者株系。
此外,作者还预测了一些有趣的生态动力学模式:当三个部分生产者各自产生一种铁载体,并且每个菌株都能利用下一个菌株产生的铁载体时,可能形成“剪刀石头布”式的循环关系并导致系统振荡(图3D,5)。这种非线性现象对维持生物多样性具有重要意义。此外,当两个菌株互相盗窃对方的铁载体时,在适当的盗窃强度下可能实现共存,这为盗窃行为促进生物多样性提供了新的思路。
综上所述,铁载体相作有成为微生物博弈理论的理想实验平台的潜力。
图3. 铁载体介导的互作可以体现多样的生态互作模式。A. 铁获取的三种策略类型,通过每个微生物产生和吸收何种铁载体来进行区分,其功能群在下方用示意图表示。带边缘的“自受体”(self-recognizer)和无边缘的“盗受体”(cheating-recognizer)用旗形来分别表示。带叉的方框表示"不存在"。本图中所有铁载体介导的互作使用与图1D相同的符号系统,微生物边缘的颜色表示不同的策略类型(绿色:纯生产者;橙色:部分生产者;红色:纯盗窃者)。B. 铁载体互作矩阵(SIM)描述了两个微生物之间由铁载体介导的互作关系。矩阵中的每个元素表示两个菌株的自受体和盗受体是否在类型上重叠(用1和0值表示),或缺失该类识别受体(用NA值表示)。‘任意值Any Value表示识别受体的重叠与否都可接受。C. SIM矩阵不同元素所表征的互作关系示例,包括趋同生产(左上,SIM(1,1)=1)、铁隔离(右上,SIM(1,1)=0)、盗窃(左下,SIM(1,2)=1或SIM(2,1)=1)和盗窃者竞争(右下,SIM(2,2)=1)。D. 铁载体介导的互作可以实现的经典生态互作场景,如竞争排除、公地悲剧、部分盗窃和互相盗窃(从左列到右列,D(1)到D(4))。每一列从上到下展示了菌株的产生和利用关系、对应的SIM矩阵、简化的网络表示(从菌株i到j的箭头表示菌株i产生的铁载体可被菌株j利用)以及动力学模型的示意相图。此外,三菌株系统可能展现“剪刀石头布”式的动力学关系,这也在D(5)中展示。
4. 为多样性而战——次级代谢通过化学创新的进化意义
在深入理解铁载体网络的复杂性之后,一个根本性的问题浮现出来:微生物为什么会进化出如此多样的铁载体结构,而不是简单地趋同于最有效的一种?作者从分子机制、演化驱动、和复杂系统理论三个层面探讨了这一问题。
在分子机制的层面上,NRPS(非核糖体肽合成酶)的模块化结构为铁载体多样性的产生提供了理想平台。这种“流水线”式的工作方式允许通过模块重排和突变产生新的铁载体结构。特别是,其模块中的保守位点使重排成为可能,而选择底物的A domain的突变则可以改变底物特异性。这解释了为什么NRPS途径是微生物中最丰富的次级代谢途径之一,不仅用于合成铁载体,还用于产生抗生素等多种竞争性次级代谢产物(图4A),而对这种多样性产生的分子机制的理解,有助于合成生物学理性设计新的基因簇。
关于演化的驱动力,作者探讨了几种可能的解释。最简单的假设是中性漂变,即新的铁载体类型在没有选择压力的情况下随机产生。然而,其他资源获取途径相对保守的现象表明,铁载体系统可能存在特殊的选择压力。而另一种解释是,铁载体多样化有助于生产者逃避偷盗者。即使铁载体结构的微小变化也可能显著改变其识别特异性,帮助生产者避免被其他微生物偷盗(图4B)。还有一种解释认为,噬菌体的选择压力会促使识别受体发生变化,进而导致铁载体结构的相应改变,因为许多噬菌体利用铁载体受体作为入侵细胞的途径(图4C)。此外,宿主-病原体的互作也可能推动铁载体的多样化,比如细菌进化出新的铁载体结构来逃避宿主免疫系统的识别。
从更广泛的理论视角来看,铁载体多样化展示了生命系统中"化学创新"的重要性。正如斯图尔特·考夫曼(Stuart Kauffman)所述,创新往往发生在生命探索其当前可能性边界的过程中。微生物不仅消耗现有资源,还不断创造新的化学物质,扩展了它们的生存空间。新代谢产物的出现呈指数级扩大了化学空间,催生新的互作关系,可能引发群落的相变。随着化学维度的增加,竞争原理可能不再限制生物多样性,因为生命本身在不断推动边界的扩展(图4D)。
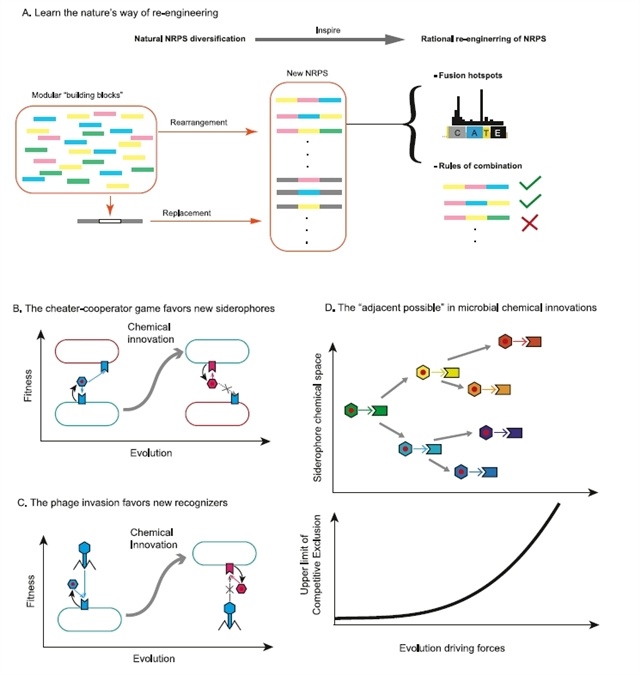
图4. 铁载体多样化的进化和分子基础及其意义。A. 铁载体多样化如何启发NRPS的理性重编程。自然界中NRPS的多样化过程,如模块化部件的重排和替换,启发了NRPS的理性合计,通过理解重组热点和组合规则来创造新的产物。B. 生产者-偷盗者的博弈如何促进新铁载体的产生。上图中展示了偷盗者和生产者之间的博弈如何驱动新铁载体锁钥对的出现。C. 噬菌体选择压力如何促进新识别受体的形成。图中展示了噬菌体入侵如何驱动识别受体的改变和新铁载体锁钥对的形成。D. 微生物化学创新中的"邻近可能性"概念。上图展示了一个已有的铁载体锁钥对(绿色类型)如何在某些进化驱动力下扩展到其邻近可能空间(不同颜色)。下图展示了这种铁载体多样化如何提升竞争排除定律的上限。
5. 讨论
作者在讨论部分首先强调了"铁网"构建的实践意义。微生物群落对人类健康、农业和工业都至关重要,但由于其复杂性,研究者们一直缺乏精确调控的手段。虽然铁互作只是微生物间众多互作方式之一,但铁元素对几乎所有微生物的生长都必不可少,这使得基于铁载体的调控具有普适性。更重要的是,铁载体与识别受体之间的特异性为精确干预提供了可能,而智能化的铁分型方法则可以利用不断增加的序列数据,通过计算方法去推断这些互作关系。
从理论价值来看,次级代谢主动向新化合物演化,以扩充其生存的可能性空间,这将生态学也引入一个新的领域——几乎所有生命形式都在创造新的生存策略,从微生物的抗生素军备竞赛到人类的创新。传统生态理论主要基于预定义的化学维度,而"邻近可能"的概念仍然停留在理论层面。研究微生物如何扩展其可能性空间,特别是通过次级代谢,可能为生态理论提供新的视角。
然而,这个框架也存在重要的局限性。首先,作者简化了“调控”的存在。“从序列到生态”的方法就假设了:基因组中合成酶或受体基因的存在性就意味着其发挥功能。实际上,相关基因的表达受到铁可利用性、种群密度和竞争者存在等多个因素的调控。其次,铁介导的互作与更广泛的代谢过程相关联,不同的营养限制可能影响铁载体和受体的表达。
而对于发展铁网络这个未来大方向,仍有许多谜题待解。例如:为什么不同铁载体的使用范围如此不同?为什么没有进化出能够吸收所有铁载体的“超级受体”?细胞如何根据不同环境和竞争者调节其铁策略?我们能否开发出构建铁网的通用算法?革兰氏阳性菌和阴性菌之间如何交流?这些问题的解答需要分子结构、生物信息学、生态建模和医农实验等多个领域的通力合作。
总的来说,“铁网”的构建将是一个宏大的跨学科项目。未来,作者期待它不仅能帮助我们理解和调控微生物群落,还可能为生命系统的创新和多样性提供新的理论视角。作者希望各领域的研究者共同参与这一努力。
QB期刊介绍
Quantitative Biology (QB)期刊是由清华大学、北京大学、高教出版社联合创办的全英文学术期刊。QB主要刊登生物信息学、计算生物学、系统生物学、理论生物学和合成生物学的最新研究成果和前沿进展,并为生命科学与计算机、数学、物理等交叉研究领域打造一个学术水平高、可读性强、具有全球影响力的交叉学科期刊品牌。
《前沿》系列英文学术期刊
由教育部主管、高等教育出版社主办的《前沿》(Frontiers)系列英文学术期刊,于2006年正式创刊,以网络版和印刷版向全球发行。系列期刊包括基础科学、生命科学、工程技术和人文社会科学四个主题,是我国覆盖学科最广泛的英文学术期刊群,其中12种被SCI收录,其他也被A&HCI、Ei、MEDLINE或相应学科国际权威检索系统收录,具有一定的国际学术影响力。系列期刊采用在线优先出版方式,保证文章以最快速度发表。
中国学术前沿期刊网
http://journal.hep.com.cn

特别声明:本文转载仅仅是出于传播信息的需要,并不意味着代表本网站观点或证实其内容的真实性;如其他媒体、网站或个人从本网站转载使用,须保留本网站注明的“来源”,并自负版权等法律责任;作者如果不希望被转载或者联系转载稿费等事宜,请与我们接洽。